





In this commentary, James Umen explores the findings of Signorovitch et al., in this issue of GENETICS, who discovered an unusual form of sexual exchange within a population of bdelloid rotifers, a group previously thought to have evolved without sex for many millions of years.
FIFTEEN years ago, Mark Welch and Meselson published a landmark paper that threw down a gauntlet to evolutionary biologists. They presented the first compelling molecular evidence that a large group of animals called bdelloid rotifers had been evolving for many millions of years without sex (Mark Welch and Meselson 2000). Their findings challenged the importance of sexual reproduction for long-term maintenance of a eukaryotic lineage and added considerable weight to the notion of asexual bdelloids as an evolutionary “scandal” (Smith 1986). Subsequent work by Meselson, Mark Welch, and colleagues uncovered an alternative explanation for their original findings based on the discovery that bdelloids are ancient tetraploids. Copies of genes that were originally interpreted as deeply diverged homologs that had ceased recombination were actually found to be homeologs that had most likely arisen from genome hybridization (Mark Welch et al. 2008). However, this finding did not rule out asexual evolution in bdelloids, in which among hundreds of cultured samples and natural populations, male bdelloids have not been observed (Birky 2010). Moreover, the case against sex in bdelloids received further support in a recent publication describing the sequencing of a bdelloid genome whose structure is incompatible with conventional meiosis (Flot et al. 2013). Instead of typical sets of homologous chromosomes, the tetraploid bdelloid genome is full of rearrangements and translocations that place some genes that were once allelic pairs on the same chromosome. Case closed for asexual evolution in bdelloids? Apparently not. In this issue of GENETICS, Meselson and co-workers report the surprising finding that bdelloids have a sex life after all—though clearly not a conventional one (Signorovitch et al. 2015).
It is worth revisiting the reason why apparent loss of sexuality in bdelloids has been a conundrum for evolutionary biologists [though see Birky (2010) for an alternative view]. Sexual reproduction via meiosis (recombination and genome reduction) and syngamy (gamete fusion) is an ancient eukaryotic innovation (Goodenough and Heitman 2014), and its maintenance and near ubiquity speak to its importance. There are many reasons why sexual reproduction is thought to be beneficial and remains under long-term selection, not least of which are the efficiency with which deleterious alleles can be purged from a sexual population and the rapidity with which an adaptive landscape can be explored through the generation of sexual recombinants. Lineages that lose sex tend to adapt more slowly than sexual ones, and they may undergo genomic degeneration through the slow accumulation and fixation of deleterious mutations, a process known as Muller’s ratchet. In the short term, however, asexual (i.e., clonal) reproduction can be a highly successful means of propagating and expanding a population with a well-adapted genotype because clonal progeny do not suffer the cost of genomic reshuffling or the twofold cost of males that are associated with sex (Butlin 2002; Otto 2009). In the face of these opposing costs and benefits, eukaryotes have evolved a diverse range of reproductive strategies including obligately sexual (e.g., humans and many other animals), facultatively sexual (e.g., algae, fungi, and many unicellular eukaryotes), and completely asexual (e.g., dandelions). What distinguishes complete asexuals from the first two groups is that they tend to be short-lived twigs on the tree of life and are usually embedded within closely related sister taxa that have retained sex (Butlin 2002; Simon et al. 2003). The interpretation of this short, twiggy distribution is that asexual species may arise fairly often and can flourish transiently, but without some form of sex and recombination, their long-term prospects are bleak. Flying in the face of this idea were bdelloid rotifers, a class of at least 20 genera and 460 species that seems to have done just fine for tens of millions of years without sexual reproduction compared to related taxa such as monogonont rotifers that have retained facultative sex (Segers 2007; Mark Welch et al. 2009).
In their recent study, Meselson and co-workers took a new approach to investigating the potential for sex in bdelloids (Signorovitch et al., 2015). They looked at population data from different geographic isolates within the same bdelloid species, Macrotrachela quadricornifera, in order to uncover possible evidence of genetic exchange. They further narrowed their search by focusing on a set of isolates from different locations that shared highly similar mitochondrial DNA, reasoning that if there were genetic exchange, it would be most apparent in isolates with recently shared maternal ancestry. They examined sequences from four loci that had previously been studied and were known to be physically unlinked in related species (Mark Welch et al. 2004).
As mentioned earlier, bdelloids are ancient tetraploids and therefore have two highly diverged homeologs for many genes. Signorovitch et al. (2015) focused on two pairs of homeologs and their surrounding regions (hisA, hisB, hsp82A, and hsp82B). For each locus, there are two copies per genome (e.g., hsp82A1 and hsp82A2) that were formerly allelic pairs but, as a result of genome rearrangements, may no longer recombine. By examining sequence similarity patterns at these four loci in a set of independent geographic isolates of M. quadricornifera, Signorovitch et al. (2015) were able to infer that some form of sexual genetic exchange was taking place.
Before describing the pattern of allele sharing they found in their isolates, it helps to first consider the expected observations in the case where no sexual exchanges occur. If we had four isolates (designated w, x, y, and z), in the absence of sex, we would expect that all the hsp82A1 sequences from these isolates (e.g., hsp82A1.w, hsp82A1.x, hsp82A1.y, and hsp82A1.z) would cluster in a single well-defined clade and that all the hsp82A2 sequences from each isolate would cluster in another well-defined clade (Figure 1A). Furthermore, the topology of the hsp82A1 and hsp82A2 subclades should be congruent because all alleles in all isolates have been evolving independently of each other for exactly the same number of generations (Figure 1A). Instead, Signorovitch et al. (2015) observed instances where, for example, the hsp82A1.w-hsp82A1.z distance was nearly zero, whereas the hsp82A2.w-hsp82A.z distance was much larger (Figure 1B). Gene conversion is one way in which genetic information can be exchanged and lead to loss of interallelic diversity (Figure 1B), but the similarity pattern caused by gene conversion within an isolate is different from the pattern of allele sharing between isolates that Signoravitch et al. observed (Figures 1, B and C). A nonsexual explanation for allele sharing could be horizontal gene transfer (HGT), in which a copy of the hsp82A1 region from one isolate replaced that of another (because no extra copies were found for genes in any of the loci examined an HGT-based explanation also would have to invoke an allelic replacement and not addition of a new sequence elsewhere). Arguing strongly against HGT were the findings of identical allele-sharing patterns at four independent loci (hsp82A, hsp82B, hisA, and hisB), where in all cases one of the two alleles at the locus showed a near-zero distance between isolates w and z, whereas the other allele showed a much larger distance between the same two isolates (Figure 1D). In other words, there appeared to have been a genome-wide exchange of genetic information between distinct bdelloid isolates, a hallmark of sex.
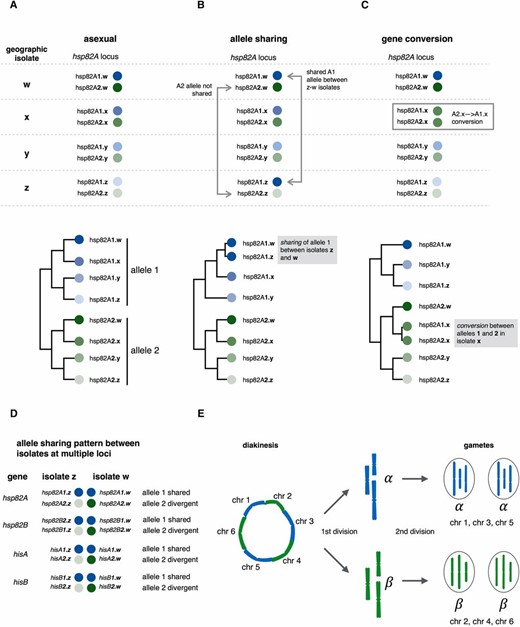
Evidence for sexual exchange in bdelloids from patterns of allele sharing and a proposed meiotic mechanism. (A–C) Three scenarios for patterns of allelic similarity among geographic isolates of the bdelloid species M. quadricornifera at the hsp82A locus. The upper portion depicts similarity patterns among alleles. The lower portion depicts the phylogenetic relationships between them. Individual isolates are designated by letters w, x, y, or z. (A) Upper panel. Each isolate has two alleles (hsp82A1 or hsp82A2) shaded blue and green, respectively, with degree of shading representing relative similarity between the same allele in different geographic isolates. Lower panel. Phylogeny expected with asexual evolution in the population. Alleles 1 and 2 evolve independently, and the divergence patterns for each are congruent. (B) Upper panel. Recent genetic exchange between the w and z isolates for hsp82A1 (but not hsp82A2) leads to near identical sequences between z and w isolates. Lower panel. hsp82A phylogeny showing incongruence between hsp82A1 and hsp82A2 allele histories reflected by different patterns in the lower and upper portions of the tree. (C) Upper panel. Recent gene conversion between the hsp82A1 and hsp82A2 alleles within isolate x. Lower panel. The phylogenetic incongruence generated by gene conversion within an isolate is different from that caused by allele sharing between isolates as shown in B. (D) Similarity of allele-sharing patterns at four independent loci in M. quadricornifera (hsp82A, hsp82B, hisA, and hisB) where in each case the w and z isolates are nearly identical for one allele but divergent for the other. (E) Simplified diagram of atypical meiosis seen in Oenothera species with permanent translocation heterozygosity that may be similar to meiosis in bdelloids. Six chromosomes are shown (chr 1–chr 6). Chromosome pairing occurs only at ends and results in ring formation at diakinesis with alternating parental duplexes shaded blue and green. At meiosis I, alternating chromosomes cosegregate into two complexes (α and β), each containing half the parental chromosomes with no recombination between genic regions. The union of an α and a β gamete restores a full set of chromosomes to an F1 progeny.
The observation of similar allele-sharing patterns at all four independent loci examined by Signorovitch et al. (2015) was not expected for a population undergoing standard meiotic sex, where each unlinked locus should exchange independently, but we already know that standard meiosis is incompatible with bdelloid genome structure (Flot et al. 2013). Signorovitch et al. (2015) instead speculate that bdelloids evolved a meiotic mechanism that resembles what is found in an unusual group of flowering plants from the genus Oenothera (evening primrose). Like bdelloids, Oenothera genomes from some species are filled with translocations that prevent normal meiotic pairing and recombination. Instead, the Oenothera chromosomes have evolved a variant type of premeiotic interaction in which ordered chromosome rings form via limited pairing at homologous end sequences (Golczyk et al. 2014)(Figure 1E).
Importantly, owing to a stereotyped alternating order of parental chromosomes, the gametes generated in Oenothera meiosis are not random mixtures but instead consist of two types (termed α and β)(Figure 1E). Although α/α and β/β offspring are nonviable owing to genetic imbalances, α/β offspring contain a full complement of genetic information and are viable and fertile. If a similar mechanism were operating in bdelloids, it would explain how allelic patterns could be shared genome-wide across isolates because hsp82A1, hsp82B1, hisA1, and hisB1 and their respective chromosomes would cosegregate as a unit into one type of gamete, whereas hsp82A2, hsp82B2, hisA2, and hisB2 would cosegregate into the alternative gamete type. Although this segregation mechanism could incur a 50% viability penalty due to nonviable α/α or β/β pairings in half the offspring, the penalty could be greatly offset by the benefits of hybridization between diverged genomes in α/β offspring.
Though an Oenothera-like meiotic mechanism in bdelloids is plausible and even likely, it remains a speculative model that must be verified. In addition, few population genetic models for Oenothera-like translocation hybrids exist and must operate with assumptions and simplifications that may or may not be correct (Holsinger and Ellstrand 1984). Nonetheless, even a limited means of genetic exchange—as observed now in bdelloids—may be enough to keep the population fit and thriving without classic meiotic recombination. Unfortunately, sex in bdelloids is still extremely rare, and without a means of directly observing sexual development, the meiotic mechanisms used by bdelloids are likely to remain hidden. The scandalous poster child for asexual evolution has become less of a scandal but no less a mystery.
Acknowledgments
The author's laboratory work is supported by National Institutes of Health grants R01GM078376 and R01GM092744.
Footnotes
Communicating editor: J. Heitman

{kind=link}