




Abstract
In recent years, the synergistic relationship between NADPH oxidase (NOX)/dual oxidase (DUOX) enzymes and peroxidases has received increased attention. Peroxidases utilize NOX/DUOX-generated H2O2 for a myriad of functions including, but not limited to, thyroid hormone biosynthesis, cross-linking extracellular matrices (ECM), and immune defense. We postulated that one or more peroxidases produced by Caenorhabditis elegans would act in host defense, possibly in conjunction with BLI-3, the only NOX/DUOX enzyme encoded by the genome that is expressed. Animals exposed to RNA interference (RNAi) of the putative peroxidase genes were screened for susceptibility to the human pathogen Enterococcus faecalis. One of three genes identified, skpo-1 (ShkT-containing peroxidase), was studied in depth. Animals mutant for this gene were significantly more susceptible to E. faecalis, but not Pseudomonas aeruginosa. A slight decrease in longevity was also observed. The skpo-1 mutant animals had a dumpy phenotype of incomplete penetrance; half the animals displayed a dumpy phenotype ranging from slight to severe, and half were morphologically wild type. The SKPO-1 protein contains the critical catalytic residues necessary for peroxidase activity, and in a whole animal assay, more H2O2 was detected from the mutant compared to the wild type, consistent with the loss of an H2O2 sink. By using tissue-specific skpo-1 RNAi and immunohistochemical localization with an anti-SKPO-1 antibody, it was determined that the peroxidase is functionally and physically present in the hypodermis. In conclusion, these results characterize a peroxidase that functions protectively in the hypodermis during exposure to E. faecalis.
HEME-containing peroxidases play critical, wide-ranging roles in biological systems. Once called the “animal heme peroxidases,” members of the peroxidase–cyclooxgenase superfamily are actually found in all kingdoms of life (Zamocky et al. 2008). The roles of most are poorly characterized, but some are clearly involved in immune defense. For example, the most famous and best-studied member of this group, myeloperoxidase (MPO), is found in the granulocytes of neutrophils where it catalyzes the formation of the potent oxidant HOCl from H2O2 and Cl− to kill invading microbes (reviewed by Klebanoff 2005). Another is lactoperoxidase (LPO), which is found on mucosal surfaces and generates the protective oxidant hypothiocyanite (OSCN−) from H2O2 and thiocyanate (SCN−). This process is impaired in the lungs of patients with cystic fibrosis (CF), contributing to the poor clearance of pathogens such as Pseudomonas aeruginosa and Staphylococcus aureus, the most common causes of lung infection in patients with CF (Conner et al. 2002; Geiszt et al. 2003; Forteza et al. 2005; Moskwa et al. 2007).
Many of these peroxidases are functionally associated with members of the NADPH oxidase (NOX)/dual oxidase (DUOX) family of proteins. These enzymes generate the H2O2 required as substrate for the peroxidases. For example, Nox2 is the source of H2O2 for MPO. DUOXs differ from NOXs in that they encode a peroxidase domain in addition to the oxidant-generating NADPH oxidase domain (reviewed by Rada et al. 2008; Sumimoto 2008). Despite the fact that they have a peroxidase domain, DUOXs appear to still associate with separate peroxidases. For example, LPO utilizes H2O2 generated by Duox1 or Duox2 (Conner et al. 2002; Geiszt et al. 2003; Forteza et al. 2005). In fact, the peroxidase domains of human Duox1/2 are reported to lack peroxidase activity and may have other functions, such as serving as an interaction domain for separate, active peroxidases (Meitzler and Ortiz de Montellano 2009, 2011; Meitzler et al. 2013).
Caenorhabditis elegans has been used as a model host to study various aspects of the innate immune response, including the purposeful generation of reactive oxygen species (ROS) as a defense mechanism (Chavez et al. 2007, 2009). The animal encodes only one functional NADPH oxidase, a dual oxidase called BLI-3 (Edens et al. 2001). When exposed to human pathogens such as the Gram-positive, opportunistic bacterium, Enterococcus faecalis, H2O2 is released (Chavez et al. 2007). The response appears protective, as its loss by reducing the expression of bli-3 by RNA interference (RNAi) renders the animals more sensitive to killing by the pathogen (Chavez et al. 2009). Using indirect immunofluorescence, BLI-3 was localized to the hypodermis, which is essentially the “skin” of C. elegans (Edens et al. 2001). There is also speculation that BLI-3 may be present in the intestinal cells (Chavez et al. 2007, 2009). In addition to playing a role in innate immunity, BLI-3 is essential to the normal development of the worm because it contributes to the generation of the tyrosine-linked collagen necessary for proper biogenesis of the cuticle, i.e., the exoskeleton of the animal. Its role appears twofold. First, it generates the necessary substrate H2O2, and then it oxidizes the tyrosines of the collagen proteins that then combine to form di- and trityrosines, thereby cross-linking the cuticle (Edens et al. 2001). Unlike human Duox1/2, the peroxidase domain of BLI-3 has low levels of peroxidase activity that is essential to this process, and mutations in the peroxidase domain that disrupt this activity result in a “blistered” (bli) phenotype (Brenner 1974; Simmer et al. 2003; Meitzler and Ortiz de Montellano 2009, 2010, 2011). However, more recent work has demonstrated that the peroxidase domain of BLI-3 is not the only peroxidase involved in this process. A separate peroxidase, MLT-7, also contributes to cuticle cross-linking and loss of this activity results in the same bli phenotype as loss of the BLI-3 peroxidase domain. Additionally, loss of both peroxidase activities greatly increases the severity of cuticle blistering (Thein et al. 2009).
Because of the prevalent involvement of peroxidases in immune responses, we hypothesized that C. elegans might also utilize a peroxidase(s) in host defense, perhaps in conjunction with its NADPH oxidase, BLI-3. The results of an earlier investigation indicated that the peroxidase domain of BLI-3 is not involved, as point mutants in this domain had wild-type resistance to E. faecalis, despite their blistered phenotype (Chavez et al. 2009). In this study, we characterized the effects of reducing the expression of putative peroxidases encoded by the C. elegans genome on susceptibility to E. faecalis. We identified three genes whose loss resulted in susceptibility. One gene, named skpo-1, for ShkT-containing peroxidase, was investigated in depth.
Materials and Methods
Creation of complementary DNA pL4440 constructs for double-stranded RNA production in Escherichia coli
A standard RNA isolation procedure was used to procure total RNA from healthy N2 adults grown on Escherichia coliOP50. We followed the first-strand complementary DNA (cDNA) synthesis protocol from the Life Technologies SuperScript III Synthesis system to produce first-strand total cDNA. We then followed a standard PCR protocol using primers specific to F32A52.A, F32A52.B, and pxn-1, respectively (Supporting Information, Table S5). PCR products were digested and ligated into pL4440 and transformed into chemically competent E. coliHT115 to produce double-stranded RNA (dsRNA) to the gene of interest, respectively.
C. elegans strains and growth conditions
C. elegans strains were grown and maintained as previously described (Hope 1999). The strains used for the hypodermal and intestinal RNAi studies are as follows: rde-1(ne219); Is[wrt-2prom::RDE-1::unc54 3′utr; myo2p::RFP3] and sid-1(qt9); Is[vha-6::sid-1], respectively. The RB1437 [skpo-1 (ok1640) II] strain is a partial deletion mutant for skpo-1 and was verified via sequencing (www.wormbase.org). The RB1437 [skpo1 (ok1640) II] strain was created by the C. elegans Reverse Genetics Core Facility at the University of British Columbia, part of the International C. elegans Gene Knockout Consortium and obtained through the Caenorhabditis Genetics Center. The independent lines of skpo-1 strains, GF89–GF91, were obtained through backcrossing the RB1437 [skpo-1 (ok1640) II] strain with the wild-type N2 Bristol strain five times (backcrossing, BC ×5).
RNAi
RNAi was performed by exposing L1–L4 stage larvae to E. coliHT115-expressing dsRNA to target genes. RNAi clones were obtained from the C. elegans library (Fraser et al. 2000; Kamath et al. 2003). skpo-1 worms exposed to pathogens were prone to a maternal bagging phenotype; therefore, cdc-25.1 RNAi was used to induce sterility in some experiments (van der Hoeven et al. 2011).
Survival and longevity assays
Unless otherwise indicated, the following bacterial strains were used: OP50 (E. coli) (Brenner 1974), OG1RF (E. faecalis) (Dunny et al. 1978), and PA14 (P. aeruginosa) (Rahme et al. 1995). Exposure to RNAi, survival assays, and longevity assays were performed as previously described (Garsin et al. 2001, 2003; Kim et al. 2002; van der Hoeven et al. 2011). Briefly, for E. faecalis survival assays, E. faecalis grown in brain heart infusion (BHI) medium for 5 hr was seeded (10 μl) onto BHI plates (gentamycin 50 μg/ml) and incubated at 37° for 24 hr, whereas for P. aeruginosa survival assays, P. aeruginosa was cultured in Luria broth (LB) overnight at 37°, seeded (10 μl) onto slow-killing plates, and incubated first for 24 hr at 37° and then for the duration of the experiment at 25°. For the E. coli longevity assays, 20× E. coli was seeded (100 μl) onto nematode growth (NG) medium plates supplemented with 5-Fluoro-2′-deoxyuridine (25 μg/ml) and streptomycin (25 μg/ml). Seeded NG plates were incubated at 25° for 24 hr. In all assays, a total of 90 L4 larvae were transferred to three replica plates and the assay was performed at 25°. Worms were scored as live and dead at various time points.
Amplex Red assay for H2O2 measurements
The Amplex Red hydrogen peroxide/peroxidase kit (Invitrogen Molecular Probes, Eugene, OR) was previously adapted to C. elegans to measure pathogen-stimulated hydrogen peroxide release (Chavez et al. 2007, 2009). The same protocol was followed with the following modifications: L4 worms were exposed to a bacterial strain for 12 hr, and the fluorescence of 30 worms per well was measured after 30 min of incubation with Amplex Red (540/590 nm excitation and emission, respectively). A total of 80 μM diphenyleneiodinium chloride (DPI) (TCI, Tokyo) was added to some wells and allowed to incubate for 15 min prior to addition of Amplex Red, HRP, and Na2PO4 (pH 7.4), and fluorescence was measured as above.
Indirect immunofluorescence
Immunofluorescence for C. elegans was performed according to Seydoux and Dunn (1997) on young adult animals. Custom peptide synthesis, rabbit immunization, and affinity purification of the SKPO-1 polyclonal antibody was performed by LifeTein (South Plainfield, NJ). The CRVGRRAFDIENGSR peptide corresponds to the C terminus of the C. elegans protein SKPO-1. Adult N2 and GF89 worms were imaged using an Olympus IX81 automated inverted microscope and Slidebook (version 5.0) software along with the SKPO-1 polyclonal primary antibody and Alexa Fluor 488 goat antirabbit IgG secondary antibody to localize SKPO-1 in C. elegans.
Immunogold labeling and transmission electron microscopy
Approximately 1000 N2 and GF89 animals were raised under standard conditions to the young adult stage. They were rinsed off of the propagation plates with M9 and spun down at 2000 rpm for 1 min. The worms were sequentially resuspended and pelleted three times in 1 ml M9 (Hope 1999). After the final wash, they were resuspended in 1 ml 3% formalin, 0.15% glutaraldehyde in Millonig’s buffer, pH 7.45, and fixed for 3 days at 40°. Then the buffer was drawn off and the samples were incubated in 0.1% fresh made sodium borohydride in Millonig’s buffer (room temperature, RT, for 10 min). This was followed by incubations in Millonig’s buffer (RT, 10 min, repeated twice). The samples were then dehydrated at room temperature by incubating in 50% ethanol (10 min, twice) and 70% ethanol, (15 min, twice). The samples were then permeated with 50% LR White resin and 50% ethanol for 60 min, followed by an overnight incubation at room temperature in 50% LR White resin sealed on a rotator. The samples were then incubated at room temperature for 4 hr in 70% LR White resin and then for 5 hr in 100% LR White resin, while on a rotator. The pellets were then sealed into BEEM capsules and allowed to polymerize in a 53° oven overnight. Thin sections (120 nm) of the LR White blocks were cut using a DiATOME diamond knife and a Leica Ultracut R microtome. Sections were floated onto 200 mesh nickel grids. The grids were then floated in 50 mM glycine (RT, 15 min) and then washed twice by floating in drops of PBS. The samples were blocked by floating them for 30 min in Aurion Blocking Solution followed by washes in incubation buffer (PBS with 0.1% Aurion BSA-c) (5 min, three times). The SKPO-1 antibody was diluted into the incubation buffer at 1:1000 and the grids were incubated for 1.5 hr at RT. This was followed by washes in incubation buffer (10 min, four times) and then incubation with the secondary antibody (EMS goat antirabbit IgG with 10 nm gold) diluted 1:20 in incubation buffer. The samples were washed again in incubation buffer (10 min, five times) followed by PBS (10 min, three times). A postfix treatment of 3% glutaraldehyde was applied for 10 min followed by a final wash with distilled water (5 min, two times). The grids were dried in a 70° oven and then imaged using a JEOL 1200 transmission electron microscope at 60 kV and captured with a 2k × 2k Gatan CCD camera.
Statistical analysis
Amplex Red, survival, and longevity assays were analyzed using GraphPad Prism version 5.0 (GraphPad Software, San Diego). Student’s paired t-test was used to determine the statistical significance of the Amplex Red data. P-values of <0.05 were considered to be statistically significant. Statistically significant differences are denoted in the figures with asterisks. Kaplan–Meier log rank analysis was used to compare survival and longevity curves pairwise and to calculate the median survival. P-values of <0.05 were considered to be statistically significant.
Results
Host resistance to E. faecalis is affected by skpo-1
To investigate if C. elegans possesses any peroxidases involved in host defense, we utilized an RNAi-killing assay to screen candidate peroxidase genes. The candidate genes were found using the BLI-3 peroxidase domain as the BLAST query on www.wormbase.org. Twelve putative peroxidase-encoding genes were identified, 9 of which were available in the RNAi library (Fraser et al. 2000; Kamath et al. 2003), including the previously studied mlt-7 (Thein et al. 2009). Standard molecular biology techniques were used to generate the three missing RNAi constructs (Materials and Methods). Using a background in which RNAi efficiency is increased (eri-1) (Kennedy et al. 2004), the expression of these genes was reduced in C. elegans. The animals were then exposed to E. faecalis and survival was scored over time. By these means, the genes were screened for possible roles in innate immune function. The reduced expression of three genes—F09F3.5, R08F11.7, and F49E12.1—resulted in a statistically significant susceptibility phenotype (Table 1). We decided to focus on F49E12.1, as it possessed high homology to human myeloperoxidase (hMPO, 34% sequence identity) and a deletion mutant was available through the Caenorhabditis Genetics Center (CGC). As described in more detail below, the predicted protein associated with this gene contains an N-terminal metridin Shk toxin (ShkT)-like domain and a C-terminal peroxidase domain and was therefore named SKPO-1 for ShkT-containing peroxidase.
Susceptibility to E. faecalis following RNAi of putative peroxidase genes
| Gene | Susceptibility phenotype | P-value | Median survival (days) |
|---|---|---|---|
| mlt-7 control | Wild type | 0.3070 | 6 |
| 6 | |||
| pxn-1 control | Wild type | 0.9218 | 4 |
| 4 | |||
| pxn-2 control | Wild type | 0.8233 | 6 |
| 6 | |||
| C16C8.2 control | Wild type | 0.1067 | 9 |
| 9 | |||
| C46A5.4 control | Wild type | 0.7206 | 10 |
| 10 | |||
| F09F3.5 control | Susceptible | 0.0058 | 6 |
| 7 | |||
| F32A5.2a control | Wild type | 0.2499 | 5 |
| 5 | |||
| F32A5.2b control | Wild type | 0.4115 | 6 |
| 6 | |||
| F49E12.1 (skpo-1) control | Susceptible | 0.0053 | 5 |
| 9 | |||
| R08F11.7 control | Susceptible | 0.0004 | 6 |
| 7 | |||
| K10B4.1 control | Wild type | 0.7904 | 8 |
| 8 | |||
| T06D8.10 control | Wild type | 0.6636 | 8 |
| 8 |
| Gene | Susceptibility phenotype | P-value | Median survival (days) |
|---|---|---|---|
| mlt-7 control | Wild type | 0.3070 | 6 |
| 6 | |||
| pxn-1 control | Wild type | 0.9218 | 4 |
| 4 | |||
| pxn-2 control | Wild type | 0.8233 | 6 |
| 6 | |||
| C16C8.2 control | Wild type | 0.1067 | 9 |
| 9 | |||
| C46A5.4 control | Wild type | 0.7206 | 10 |
| 10 | |||
| F09F3.5 control | Susceptible | 0.0058 | 6 |
| 7 | |||
| F32A5.2a control | Wild type | 0.2499 | 5 |
| 5 | |||
| F32A5.2b control | Wild type | 0.4115 | 6 |
| 6 | |||
| F49E12.1 (skpo-1) control | Susceptible | 0.0053 | 5 |
| 9 | |||
| R08F11.7 control | Susceptible | 0.0004 | 6 |
| 7 | |||
| K10B4.1 control | Wild type | 0.7904 | 8 |
| 8 | |||
| T06D8.10 control | Wild type | 0.6636 | 8 |
| 8 |
| Gene | Susceptibility phenotype | P-value | Median survival (days) |
|---|---|---|---|
| mlt-7 control | Wild type | 0.3070 | 6 |
| 6 | |||
| pxn-1 control | Wild type | 0.9218 | 4 |
| 4 | |||
| pxn-2 control | Wild type | 0.8233 | 6 |
| 6 | |||
| C16C8.2 control | Wild type | 0.1067 | 9 |
| 9 | |||
| C46A5.4 control | Wild type | 0.7206 | 10 |
| 10 | |||
| F09F3.5 control | Susceptible | 0.0058 | 6 |
| 7 | |||
| F32A5.2a control | Wild type | 0.2499 | 5 |
| 5 | |||
| F32A5.2b control | Wild type | 0.4115 | 6 |
| 6 | |||
| F49E12.1 (skpo-1) control | Susceptible | 0.0053 | 5 |
| 9 | |||
| R08F11.7 control | Susceptible | 0.0004 | 6 |
| 7 | |||
| K10B4.1 control | Wild type | 0.7904 | 8 |
| 8 | |||
| T06D8.10 control | Wild type | 0.6636 | 8 |
| 8 |
| Gene | Susceptibility phenotype | P-value | Median survival (days) |
|---|---|---|---|
| mlt-7 control | Wild type | 0.3070 | 6 |
| 6 | |||
| pxn-1 control | Wild type | 0.9218 | 4 |
| 4 | |||
| pxn-2 control | Wild type | 0.8233 | 6 |
| 6 | |||
| C16C8.2 control | Wild type | 0.1067 | 9 |
| 9 | |||
| C46A5.4 control | Wild type | 0.7206 | 10 |
| 10 | |||
| F09F3.5 control | Susceptible | 0.0058 | 6 |
| 7 | |||
| F32A5.2a control | Wild type | 0.2499 | 5 |
| 5 | |||
| F32A5.2b control | Wild type | 0.4115 | 6 |
| 6 | |||
| F49E12.1 (skpo-1) control | Susceptible | 0.0053 | 5 |
| 9 | |||
| R08F11.7 control | Susceptible | 0.0004 | 6 |
| 7 | |||
| K10B4.1 control | Wild type | 0.7904 | 8 |
| 8 | |||
| T06D8.10 control | Wild type | 0.6636 | 8 |
| 8 |
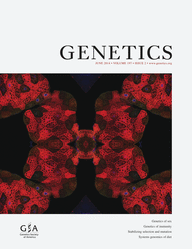
An example of the survival of skpo-1 RNAi eri-1 mutant animals on E. faecalis compared to vector control (VC) RNAi eri-1 mutant animals is shown in Figure 1A. skpo-1 RNAi animals displayed an enhanced susceptibility phenotype relative to VC RNAi (P = 0.0011). The experiment was repeated five times and the data were tabulated in Table S1. We also examined the lifespan of these animals to see if skpo-1 RNAi caused a general fitness defect. However, on E. coliOP50, lifespan of the skpo-1 RNAi animals was not significantly different when compared to VC RNAi animals (P = 0.3772) (Figure 1B and Table S2).
SKPO-1 contributes to C. elegans resistance to E. faecalis. (A) Survival of eri-1 mutant worms on E. faecalis OG1RF following exposure to vector control (VC) RNAi and skpo-1 RNAi (P = 0.0011). (B) Longevity of eri-1 mutant worms on E. coli OP50 following exposure to VC RNAi and skpo-1 RNAi (P = 0.3772). (C) Survival of wild-type and skpo-1 mutant worms on E. faecalis following exposure to cdc-25.1 RNAi (P < 0.0001). (D) Longevity of wild-type and skpo-1 mutant worms on E. coli OP50 following exposure to cdc-25.1 RNAi (P < 0.0001). (E) The relative mortality of skpo-1 mutant worms exposed to E. faecalis (EF) is expressed as a ratio of (LT50 of wild-type cdc-25.1 RNAi on EF/LT50 of skpo-1 mutant cdc-25.1 RNAi on EF) over (LT50 of wild-type cdc-25.1 RNAi on E. coli/LT50 of skpo-1 mutant cdc-25.1 RNAi on E. coli). The average of four independent experiments, with 90 animals each, was used to calculate the relative mortality. (F) Survival of wild-type and skpo-1 mutant worms on P. aeruginosa following exposure to cdc-25.1 RNAi (P = 0.3783). Error bars represent standard error of the mean (SEM) and the asterisk indicates a significant difference between wild-type and skpo-1 mutant worms (P = 0.0091). The P-values were calculated using Student’s paired t-test. The median survival for survival and longevity assays are listed in Table S1 and Table S2 along with replicates of the experiments.
To further examine the phenotype resulting from the loss of skpo-1, we obtained the partial deletion strain, RB1437 from the CGC. The deletion is between bases 827 and 3202 of the 5008 full-length transcript, which ablates a significant portion of the protein, including the critical amino acid residues necessary for peroxidase activity in classical animal heme peroxidases (reviewed by Ortiz de Montellano 2008). Strain RB1437 was backcrossed five times into our wild-type N2 strain and three lines were generated: GF89, GF90, and GF91. As shown in Figure S1, these animals were very susceptible to E. faecalis compared to wild-type N2 animals. Because significant differences in susceptibility were not observed between the strains, we continued our studies using GF89, which is referred to as skpo-1.
We observed a significant “bagging” phenotype—hatching of embryos inside the hermaphrodite that had failed to be expelled—when the skpo-1 mutant animals were exposed to E. faecalis, unlike the skpo-1 RNAi animals. There was concern that this was artificially enhancing their sensitivity to E. faecalis, thus cdc-25.1 RNAi was employed to induce sterility, as done in previous C. elegans pathogenesis studies (Shapira et al. 2006; Irazoqui et al. 2008; van der Hoeven et al. 2011) (Figure 1, C–E). skpo-1 mutant animals grown on cdc-25.1 dsRNA-expressing E. coliHT115 did not display this maternal bagging phenotype when they were subsequently exposed to E. faecalis. cdc-25.1 RNAi-exposed skpo-1 mutant worms retained a more pronounced susceptibility phenotype to E. faecalis relative to similarly exposed wild-type animals (P < 0.0001) (Figure 1C and Table S1), suggesting that the pathogen sensitivity is not completely explained by an increase in internal hatching of progeny.
Regardless of whether skpo-1 mutant worms were sterile, i.e., cdc-25.1-treated (Figure 1D and Table S2), or fecund (Figure S2 and Table S2), they displayed a slight reduction in lifespan, relative to wild type, when exposed to live E. coliOP50 (P < 0.0001, for both). Because live OP50 has been shown to have slightly pathogenic effects (Garigan et al. 2002), we also examined lifespan on heat-killed OP50 and found that cdc-25.1 RNAi-exposed skpo-1 mutant worms still displayed a reduction in lifespan (Figure S3 and Table S2). To determine if the defect in lifespan completely accounted for the pathogen sensitivity or not, the relative mortality of skpo-1 mutant worms, compared to wild type, was calculated (Figure 1F). Relative mortality is the ratio of the lethal time to kill 50% of the organisms (LT50) of pathogen-infected animals to uninfected animals with the ratio of wild-type animals normalized to one and has been used in previous work to analyze susceptibility phenotypes (Tenor et al. 2004; Chavez et al. 2007). The survival defect for the skpo-1 mutant animals on E. faecalis was more severe than on nonpathogenic E. coli compared to wild-type animals (P = 0.0091) (Figure 1E and Table S3), arguing against a general fitness defect.
To further examine the extent of the susceptibility-to-pathogen phenotype and determine if it was pathogen specific, we used P. aeruginosa instead of E. faecalis in the killing assay (P = 0.3783) (Figure 1F and Table S4). Interestingly, we observed no significant difference in survival between cdc-25.1 RNAi skpo-1 mutants and cdc-25.1 RNAi wild-type animals when exposed to P. aeruginosa. These results further argue against a general fitness defect and suggest that the function of SKPO-1 may be beneficial during infection with some, but not all pathogens.
To determine if the susceptibility phenotype on E. faecalis was a result of changes in pathogen burden, we assessed how many colony-forming units (CFUs) were in the intestines of the infected worms. Wild-type and skpo-1 mutant animals were raised on E. coliOP50 until L4 and then exposed to E. faecalis for 12 or 36 hr. The infected worms were washed to remove surface bacteria, homogenized, and then serial dilutions of the homogenates were plated. Interestingly, we observed no significant difference in CFUs per worm between the wild-type and skpo-1 mutant animals at either time point (Figure S4). The result suggests that the susceptibility phenotype of the skpo-1 mutant cannot be explained by an increased load of bacteria in the intestine.
To test for alterations in the immune response between the skpo-1 and wild-type animals a small panel of genes known to be upregulated in response to E. faecalis were examined, clec-35, -42, -60, and -71. These genes encode C-type lectins (clec), many of which are upregulated in response to pathogens (Irazoqui et al. 2010; Engelmann et al. 2011). Sterile (by exposure to cdc-25 RNAi) wild-type and skpo-1 mutant animals at the L4 stage were exposed to E. faecalis or E. coli for 18 hr, at which point the animals were lysed and RNA was extracted. By qRT-PCR, no significant difference in the expression of clec-35 and -71 was observed between the wild type and skpo-1 mutants exposed to E. faecalis. However, we observed clec-60’s expression was significantly higher in the skpo-1 mutant and there was a trend toward increased expression of clec-42, which was not statistically significant (Figure S5). These data suggest that the immune response is altered in the skpo-1 mutant.
Morphological characterization of skpo-1 mutant animals
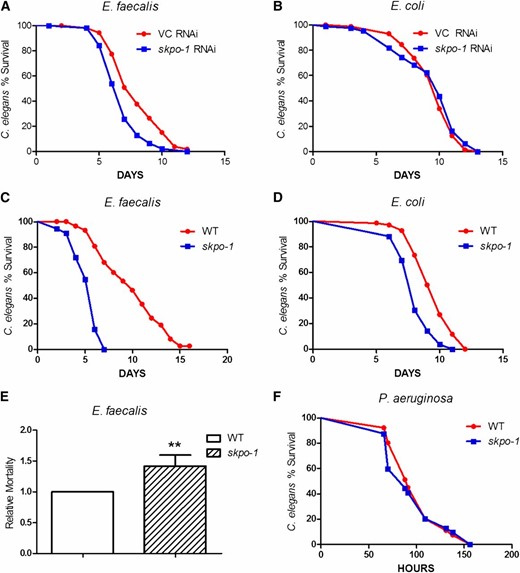
Animals deficient in the peroxidase mlt-7 displayed altered morphology, including a bli (blistered) phenotype, indicative of incomplete cross-linking of the cuticle (Thein et al. 2009). The phenotype strongly suggests that MLT-7’s peroxidase activity contributes to cuticle formation (Thein et al. 2009). Interestingly, despite this rather dramatic cuticle defect, an increase in susceptibility to E. faecalis was not observed (Table 1). To determine if SKPO-1 is also involved in cuticle biogenesis, we observed the morphology of skpo-1 RNAi and mutant animals. RNAi of skpo-1 did not result in any visible morphological change in the eri-1 mutant animals (Figure 2B). However, the skpo-1 mutant did display some morphological phenotypes. We observed young adult animals under the dissecting microscope (N = 300) and found that they ranged from very dumpy (16.5%), to slightly dumpy (32.9%), to wild type (50.6%) in appearance (Figure 2, C–E). No blistering of the cuticle was ever observed. Because a dumpy phenotype can be associated with cuticle defects (Page and Johnstone 2007), these results suggest that SKPO-1 may have some role in cuticle biogenesis, but one that is different than MLT-7. Additionally, we observed no significant difference in susceptibility between skpo-1 mutant morphotypes, i.e., all were equally hypersusceptible to E. faecalis relative to wild-type animals (Figure S1).
skpo-1 mutant displays multiple morphotypes. (A) Wild-type young adult representing the approximate average size and morphology of a typical C. elegans. (B) skpo-1 RNAi young adult raised on skpo-1 dsRNA-expressing E. coli HT115 from L1–L4 stages. (C–E) Young adult skpo-1 mutants ranged from very dumpy to wild type in size. The ×10 microscopy images are representative of >100 wild-type, skpo-1 RNAi, and skpo-1 worms observed, respectively.
SKPO-1 features and activity
As mentioned, SKPO-1 contains a ShkT-like domain at its N terminus from residues ∼21–56 (www.wormbase.org) in addition to the predicted peroxidase domain (Figure 3A). It shares this feature with MLT-7 (Thein et al. 2009), and putative peroxidases C16C8.2 and F32A5.2 (Table 1). For this reason we propose naming F49E12.1, SKPO-1, and C16C8.2 and F32A5.2, SKPO-2, and SKPO-3 for ShkT-containing peroxidase. In addition to the ShkT-like domain, the very 5′ end contains a predicted signal sequence for secretion, and cleavage is predicted to occur at Ser19 (www.predisi.de).
![Evidence that skpo-1 is a potential peroxidase. (A) Peroxidase domain sequences were aligned against the putative peroxidase domain of SKPO-1. SKPO-1 possesses the distal histidine (H222), catalytic arginine (R332), and proximal histidine (H428), which are necessary for peroxidase activity. However, SKPO-1 lacks covalent heme-binding residues (S221 and L335) that are characteristic of mammalian peroxidases (Ortiz de Montellano 2008). (B) eri-1 mutant worms were grown on VC RNAi or skpo-1 RNAi prior to exposure with either E. coli or E. faecalis for 12 hr at 25°. (C) Wild-type and skpo-1 mutant worms were grown on cdc-25.1 RNAi prior to exposure with either E. coli or E. faecalis for 12 hr at 25°. (B and C) Following exposure to E. coli or E. faecalis, the amount of H2O2 produced per minute was determined using the Amplex Red assay. Error bars represent the SEM and the asterisks indicate significant differences between eri-1 worms exposed to VC RNAi or skpo-1 RNAi that were infected with E. faecalis as well as between wild-type and skpo-1 mutant worms exposed to cdc-25.1 RNAi prior to infection with E. faecalis [P = 0.0091 (B) and P < 0.0001 (C)]. Additionally, wild-type and skpo-1 mutant worms were exposed to 80 μM diphenyleneiodinium chloride (DPI) and H2O2 levels were calculated for both E. coli- and E. faecalis-exposed animals (EC, P = 0.0752; EF, P = 0.4161, respectively) P-values were calculated via Student’s paired t-test. Data in B and C are representative of at least two independent replicates.](https://oup.silverchair-cdn.com/oup/backfile/Content_public/Journal/genetics/197/2/10.1534_genetics.113.160606/7/m_515fig3.jpeg?Expires=1716835354&Signature=bv3o-BsParp7xd3BGH5vH2iu8IN84Dq5MwhXolEs~fLBEGa1wymEwUz5MCw-3pjxFwBb45u5o7WKVVF-OeE-GBTMnzA5JH8E2QjZxNGhY5AqiWeusd4RU9ll0ofoBfVGA0xmrcBxAOzInOSIiq6Rv39Oq4EoQF7tyjWJVQtM8fMhksGTO2AQYG1UB-6o7D7FNcE-gmzibNwQ7RIW4ACHLrLCMnLEuPPc7opZeYsBASTbE3gq9sAWiUCP6ozYa42VesQN-K3Wv11aFYqZcQpi84PUN1uNWnEdCkf0sxbkRsEWpyH7xAStyslJtf74aTFC2fQuR1KR7Jp8lbXiEUCWvQ__&Key-Pair-Id=APKAIE5G5CRDK6RD3PGA)
Evidence that skpo-1 is a potential peroxidase. (A) Peroxidase domain sequences were aligned against the putative peroxidase domain of SKPO-1. SKPO-1 possesses the distal histidine (H222), catalytic arginine (R332), and proximal histidine (H428), which are necessary for peroxidase activity. However, SKPO-1 lacks covalent heme-binding residues (S221 and L335) that are characteristic of mammalian peroxidases (Ortiz de Montellano 2008). (B) eri-1 mutant worms were grown on VC RNAi or skpo-1 RNAi prior to exposure with either E. coli or E. faecalis for 12 hr at 25°. (C) Wild-type and skpo-1 mutant worms were grown on cdc-25.1 RNAi prior to exposure with either E. coli or E. faecalis for 12 hr at 25°. (B and C) Following exposure to E. coli or E. faecalis, the amount of H2O2 produced per minute was determined using the Amplex Red assay. Error bars represent the SEM and the asterisks indicate significant differences between eri-1 worms exposed to VC RNAi or skpo-1 RNAi that were infected with E. faecalis as well as between wild-type and skpo-1 mutant worms exposed to cdc-25.1 RNAi prior to infection with E. faecalis [P = 0.0091 (B) and P < 0.0001 (C)]. Additionally, wild-type and skpo-1 mutant worms were exposed to 80 μM diphenyleneiodinium chloride (DPI) and H2O2 levels were calculated for both E. coli- and E. faecalis-exposed animals (EC, P = 0.0752; EF, P = 0.4161, respectively) P-values were calculated via Student’s paired t-test. Data in B and C are representative of at least two independent replicates.
To examine SKPO-1’s peroxidase domain, we aligned it with other well-characterized peroxidase domains of interest, those contained in hMPO, BLI-3, and MLT-7. Upon alignment, we observed SKPO-1 possesses the catalytic triad (distal histidine, H222; arginine, R332; and proximal histidine, H428) necessary for peroxidase activity (reviewed by Ortiz de Montellano 2008). However, SKPO-1 lacks the residues necessary for covalent heme binding, characteristic of the animal heme peroxidase family (reviewed by Ortiz de Montellano 2008). Interestingly, human Duox1/2 (hDuox1/2) also lack the conserved aspartic and glutamic residues required for covalent linkage of the heme prosthetic group; however, both of these recombinant peroxidase domains still bind heme, albeit weakly (Meitzler and Ortiz de Montellano 2009, 2011). This noncovalent binding may explain MLT-7’s peroxidase activity even though it lacks these covalent heme-binding residues (Thein et al. 2009).
By comparison, SKPO-1 possesses all of the highlighted residues in MLT-7 (Figure 3A) and shares significant identity with hMPO (34% identity). Thus, we decided to indirectly assay SKPO-1’s potential peroxidase activity using an Amplex Red assay, modified for whole animals, that detects H2O2 (Chavez et al. 2007, 2009). Previously, we observed that exposure to E. faecalis causes a significant release in H2O2 dependent on the NADPH oxidase, BLI-3 (Chavez et al. 2007, 2009). We reasoned that loss of an important peroxidase during this release might increase the amount of H2O2 detected. After infecting VC and skpo-1 RNAi eri-1 mutant animals with E. faecalis for 12 hr, released H2O2 was measured (P = 0.0091) (Figure 3B). skpo-1 RNAi eri-1 mutant animals released significantly greater amounts of H2O2, relative to VC RNAi eri-1 mutants, consistent with the loss of a predicted H2O2 sink. The same result was observed with the skpo-1 mutants compared to wild-type animals (P < 0.0001) (Figure 3C). The difference required E. faecalis, and the concomitant release of H2O2, as no significant differences were observed between wild-type and skpo-1-deficient animals on E. coli (Figure 3, B and C).
To investigate if the increased amount of H2O2 produced by the skpo-1 mutant was dependent on BLI-3 activity, we added the NADPH oxidase inhibitor, DPI, to the assay. We previously demonstrated that BLI-3 is the only expressed NADPH oxidase encoded by the genome (Chavez et al. 2009), and DPI can be utilized to reduce its activity in wild-type animals (Chavez et al. 2007). In Figure 3C, DPI also abrogated the enhanced H2O2 generation observed in skpo-1 mutant animals. Taken together, Figure 3 supports the hypothesis that SKPO-1 is a peroxidase that utilizes H2O2 produced by BLI-3 during infection. Unfortunately, attempts to purify the protein to demonstrate definitive peroxidase activity in vitro were not successful.
SKPO-1 localizes to the hypodermis
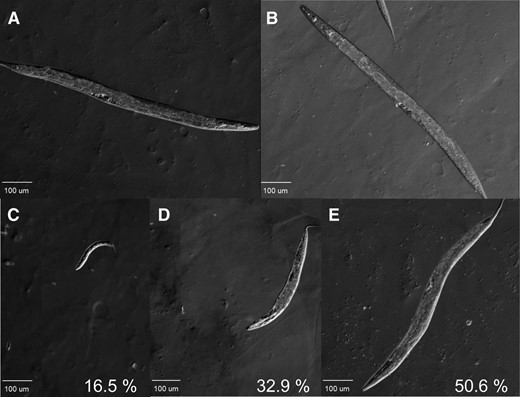
C. elegans interfaces with E. faecalis at the cuticle, synthesized by the underlying hypodermis, as it crawls through the pathogen lawn, and in its intestine, due to ingestion of the bacteria (Garsin et al. 2001). It has previously been demonstrated that C. elegans mounts immune responses at these host–pathogen boundaries, depending on the pathogen and the nature of the infection (Wong et al. 2007; Pujol et al. 2008; Irazoqui et al. 2010). To address what tissue SKPO-1’s activity is required for normal levels of resistance to E. faecalis, a tissue-specific RNAi approach was employed. RNAi-defective strains of C. elegans were used in which RNAi activity was genetically restored to specific tissues through intestinal or hypodermal-specific promoters (vha-6::SID-1 and wrt-2::RDE-1, respectively) (Melo and Ruvkun 2012). In Figure 4, A and C (and Table S1), we tested the susceptibility of the hypodermal and intestinal-specific RNAi strains to E. faecalis. As in Figure 1, these strains were exposed to VC and skpo-1 RNAi prior to infection. In the hypodermal RNAi strain, we observed an enhanced susceptibility phenotype to E. faecalis in the skpo-1 RNAi animals relative to VC RNAi (P = 0.0002); however, in the intestinal RNAi strain, no significant difference between skpo-1 and VC RNAi animals was observed when on E. faecalis (P = 0.9435). On E. coli, no significant difference was observed between VC and skpo-1 RNAi for either the hypodermal or intestinal RNAi line (P = 0.9997 and P = 0.6379; Figure 4, B and D, respectively, and Table S2). From these experiments we conclude that SKPO-1’s functional activity during pathogen exposure is required in the C. elegans hypodermis.
SKPO-1 is necessary in the hypodermis for resistance to E. faecalis infection. (A) Survival of the hypodermal RNAi strain on E. faecalis following exposure to VC RNAi or skpo-1 RNAi (P = 0.0002). (B) Longevity assay on E. coli OP50 of the hypodermal RNAi strain following exposure to VC RNAi or skpo-1 RNAi (P = 0.9997). (C) Survival of the intestinal RNAi strain on E. faecalis following exposure to VC RNAi or skpo-1 RNAi (P = 0.9435). (D) Longevity assay of the intestinal RNAi strain on E. coli OP50 following exposure to VC RNAi or skpo-1 RNAi (P = 0.6379).
To determine in which tissue SKPO-1 is produced, we used indirect immunofluorescence to visualize SKPO-1 localization in young adult animals raised under standard conditions. Using rabbits, a polyclonal primary antibody was raised against a chemically synthesized SKPO-1 peptide (see Materials and Methods). A freeze-cracking methodology was used to disrupt the cuticle of the animal and allow for internal fixation and staining (Seydoux and Dunn 1997). Following fixation, the samples were double stained with the polyclonal primary antibody to SKPO-1 and an Alexa Fluor 488 secondary antibody. Localization was clearly observed in the hypodermis of wild-type animals, and an example is shown in Figure 5, A–C and D–F (40 times and 10 times, respectively). Note the strong fluorescent staining just under the outermost layer of the animal. No internal organs showed evidence of staining, including the gonad and the intestine, which in this animal had become partially separated from the rest of the body. In contrast to wild type, skpo-1 mutant animals showed no evidence of staining in any organ (Figure 5, G–I). We also examined wild-type and skpo-1 mutant animals that had been exposed to E. faecalis for 24 hr prior to staining and did not observe any differences in the localization of SKPO-1 (data not shown).
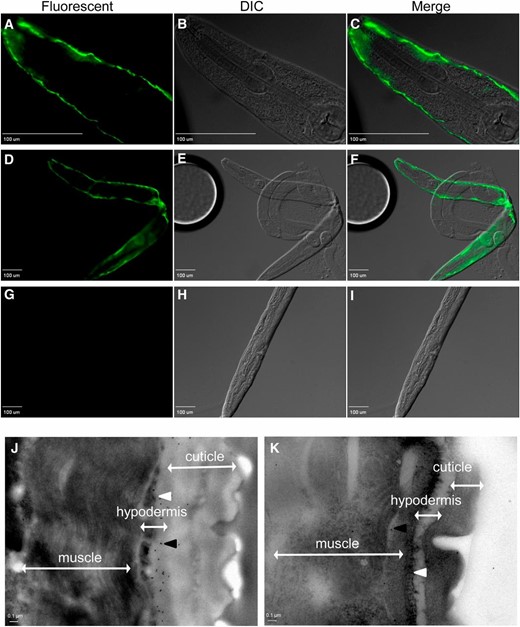
SKPO-1 localizes to the C. elegans hypodermis. (A–F and J) Wild-type and (G–I and K) skpo-1 mutant worms were immunostained with anti-SKPO-1 polyclonal antibodies and imaged using fluorescence (A–I) or transmission electron microscopy (J and K), respectively. (A–C) The ×40 magnification of a wild-type worm shows hypodermal SKPO-1 localization. (D–F) The ×10 magnification of a wild-type worm. (G–I) The ×10 magnification of a skpo-1 worm shows loss of SKPO-1 staining. (J) In wild-type worms, the black dots, indicative of immunogold labeling, are localized beneath the cuticle layer, but external to the outer hypodermal cell surface. (K) In skpo-1 mutant worms, very few black dots are observed. White arrowhead, apical hypodermal surface; black arrowhead, 10 nm gold-labeled secondary to SKPO-1). Microscopy images are representative of >100 (fluorescent) or >10 (TEM) wild-type and skpo-1 mutant worms observed.
By using a secondary antibody conjugated to 10 nm gold particles for immunogold labeling, we examined localization at higher resolution by transmission electron microscopy (TEM). As seen in Figure 5J, the black dots, indicating the gold particles, were located just under the cuticle layer in association with the hypodermal cells of wild-type animals. Hardly any particles were observed in the skpo-1 mutant animals (Figure 5K). The few observed were randomly scattered. In total, these results demonstrate SKPO-1 is both physically present and functionally active in the C. elegans hypodermis.
Discussion
In this work, we demonstrated that a previously unstudied C. elegans protein with a peroxidase domain, F49E12.1, plays a protective role during infection with E. faecalis. We named this protein SKPO-1 for ShkT-containing peroxidase, because it contains an N-terminal ShkT-like domain. The ShkT domain family was originally defined as a potassium channel blocker in the sea anemone (Metridium senile). Binding to the potassium channel requires two conserved residues that are not found in this particular ShkT-like domain of SKPO-1 or others from C. elegans (data not shown). It is postulated that the more general function of ShkT-like domains are as contact surfaces for protein interactions (Tsang et al. 2007). For this reason, SKPO-1 and other ShkT-containing peroxidases may be most closely related to the peroxidasins, subfamily 2 of the peroxidase-cyclooxygenase superfamily, and in fact, SKPO-1 was placed in this family by phylogenetic analysis (Soudi et al. 2012). These peroxidase domain-containing proteins also have protein interaction domains, but they are typically type C-like immunoglobulin domains, leucine-rich repeats, or von Willebrand factor C modules. This is in contrast to the mammalian peroxidases, MPO, LPO, and eosinophil peroxidase that lack these extra domains and belong to subfamily 1 (Zamocky et al. 2008). Additionally, it is thought that subfamily 1 evolved from subfamily 2 (Zamocky et al. 2008). Other C. elegans peroxidase domain-containing proteins that have a ShkT-like domain include MLT-7 (Thein et al. 2009), C16C8.2, and F32A5.2, but no study has yet addressed this domain’s function in the context of a peroxidase.
An unexpected finding was the discovery that SKPO-1 is located in the hypodermis and is functionally protective in this tissue against E. faecalis infection. Infection of C. elegans with E. faecalis results in colonization of the gut, leading to distension of the intestinal lumen and clear signs of physical damage, such as effacement of the microvilli (Garsin et al. 2001; Cruz et al. 2013). Though the worm is exposed to E. faecalis on its outer surface as it moves through the pathogen lawn, there is no notable colonization or characterized physical damage to the cuticle or hypodermis. In contrast, bacterial pathogens such as Microbacterium nematophilum, Xenorhabdus nematophila, and Yersinia pestis adhere to and colonize the cuticle surface (Hodgkin et al. 2000; Couillault and Ewbank 2002; Darby et al. 2002). The natural fungal pathogen Drechmeria coniospora initially adheres to the cuticle and then penetrates the hypodermis, whereas the human fungal pathogen Candida albicans first colonizes the intestine and eventually penetrates the cuticle from the inside out (Jansson et al. 1985; Breger et al. 2007). We previously showed that tissue-specific loss of bli-3 in the hypodermis also increased susceptibility of C. elegans to E. faecalis (Chavez et al. 2009). Based on the protective effects of SKPO-1 and BLI-3 in this tissue, we postulate that a hypodermal immune response does play some role in protecting C. elegans during infection with E. faecalis. The question is by what mechanism?
Several models for how these proteins might exert their protective effects can be imagined. It could be that loss of skpo-1 results in a weaker cuticle barrier that increases susceptibility. Alternatively, SKPO-1 could use H2O2 produced by BLI-3 to form more potent antioxidants, analogous to the human Duox/LPO system on mucosal surfaces (Conner et al. 2002; Geiszt et al. 2003; Forteza et al. 2005). Or the H2O2 produced by BLI-3 could act as a signaling molecule in either an autonomous or noncell-autonomous fashion. Unlike many signaling molecules, H2O2 is highly diffusible, and it could directly move into other tissues, such as the intestine, to signal responses. Interestingly, in zebrafish, H2O2 produced by a Duox in response to wounding acts as an attractant in a diffusion gradient to draw leukocytes to the area of damage (Niethammer et al. 2009). Clearly, the amount of H2O2 is important. We previously showed that loss of BLI-3 and the resulting decrease in H2O2 production caused an increase in susceptibility to infection (Chavez et al. 2009), whereas in this work we show that loss of a peroxidase and a concurrent increase in H2O2 also increases susceptibility. A similar situation is apparent from studies of infection using the model host Drosophila melanogaster (Ha et al. 2005a,b). Loss of an intestinal DUOX enzyme or an intestinal catalase both increase susceptibility to infection, but with opposite effects on ROS levels in this tissue. It is possible that SKPO-1 is catalyzing the degradation of excess H2O2 to prevent host damage. We formerly demonstrated, by a variety of means, that infection causes oxidative stress in C. elegans (Chavez et al. 2007; Mohri-Shiomi and Garsin 2008; van der Hoeven et al. 2011), much of it dependent on BLI-3 activity (van der Hoeven et al. 2011).
Interestingly, there is evidence for a hypodermal response to several pathogens that are thought to mainly cause infection in the intestine. Microarray studies that examined the transcriptional responses of C. elegans to E. faecalis, Serratia marcescens, Erwinia carotovora, Photorhabdus luminescens, S. aureus, and P. aeruginosa all noted a dramatic downregulation in the expression of genes related to cuticle biosynthesis, such as those encoding collagens (Wong et al. 2007; Irazoqui et al. 2010). The response is not thought to be due to a general reduction of gene transcription in this tissue and may be indicative of several possibilities (Wong et al. 2007). The changes in transcription of the genes could be part of a protective response that is occurring in the hypodermis to protect against pathogens. Or the changes could be purposely caused by the pathogens as part of their virulence programs to damage the host. Alternatively, the changes in expression of the cuticle biosynthetic genes could be a neutral side effect resulting from alterations in signaling due to pathogen exposure. Another microarray study observed an increase in the expression of a large number of genes related to cuticle biosynthesis in an arr-1 mutant that was found to have altered susceptibility to pathogens (Singh and Aballay 2012). arr-1 encodes for arrestin-1, a G-protein coupled receptor (GPCR) adaptor protein that is necessary for GPCR signaling in several neurons of C. elegans. Whether these changes in resistance had anything to do with alterations in the amount of cuticle biosynthetic proteins being produced was not investigated (Singh and Aballay 2012). Overall, these studies suggest that pathogen exposure, even to those pathogens not thought to directly affect the cuticle, cause major changes in the expression of the cuticle biosynthetic genes that might be indicative of a response to the infection in the hypodermis.
In conclusion, we uncovered putative peroxidases that affected susceptibility to E. faecalis in C. elegans. We focused our characterization on F49E12.1, named SKPO-1 because it contains a ShkT-like domain and a peroxidase domain. In addition to the pathogen susceptibility phenotype, loss of skpo-1 resulted in a dumpy phenotype of uneven penetrance, suggestive of some role in cuticle biosynthesis (Page and Johnstone 2007). In support of a functional peroxidase domain, we noted that SKPO-1 shares the same critical residues as MLT-7 in its active site (Thein et al. 2009), and animals mutant for skpo-1 produce significantly more H2O2 during infection. The functional and physical location for the protein was shown to be the hypodermis, which was surprising since E. faecalis has been characterized as infecting the intestine of C. elegans (Garsin et al. 2001; Cruz et al. 2013). This result, along with other evidence from the literature (Wong et al. 2007; Irazoqui et al. 2010), suggests that the hypodermis plays an important role during exposure of C. elegans to many human pathogens that do not obviously colonize or damage the cuticle, warranting further investigation.
Acknowledgments
For the TEM imaging, we thank S. Kolodziej and P. Navarro (Department of Pathology and Laboratory Medicine EM Laboratory, University of Texas Health Science Center at Houston). We thank G. Ruvkun for the tissue-specific RNAi C. elegans strains and K. McCallum, S. Arur, K. Park, T. Furata, T. Leto, A. Page, M. Cruz, and W. Nauseef for helpful input on this project. We acknowledge the Caenorhabditis Genetics Center for providing us with many of the C. elegans strains used in the study, J. Ahringer and Geneservice for the bacterial strains for RNAi, and WormBase for their help and advice in naming genes. This work was supported by Public Health Service grants R01AI076406 to D.A.G. and T32AI55449 to G.R.T. from the National Institute of Allergy and Infectious Diseases.
Footnotes
Communicating editor: B. Lazzaro
Literature Cited
Author notes
Supporting information is available online at http://www.genetics.org/lookup/suppl/doi:10.1534/genetics.113.160606/-/DC1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}